


Marchf2 녹아웃 마우스 관련 최신 연구 진전






연구자들은 Marchf2 마우스의 대립 유전자 변이를 생성하여 MARCHF2의 작용 기전을 연구했습니다. 알려진 바와 같이 많은 기념비적인 발견은 작은 시작에서 비롯됩니다. 이를 위해 유전자 연구의 훌륭한 예시인 Marchf2 녹아웃 마우스를 소개합니다.
Marchf2 유전자 녹아웃 마우스
NF-κB 필수 조절 인자(NEMO)는 세포 외 자극과 병원체 감염에 반응하는 NF-κB 및 인터페론 매개 신호 전달 과정에서 기능하는 핵심 조절 단백질입니다. NEMO의 엄격한 조절은 숙주 선천성 면역 반응과 항상성 유지에 필수적입니다. 연구자들은 E3 리가아제 MARCHF2가 세균 또는 바이러스 감염 시 NEMO 매개 신호 전달의 새로운 음성 조절 인자임을 다음과 같이 보고했습니다. MARCHF2는 감염 후기 단계에서 NEMO와 직접 상호작용하고 NEMO의 Lys326 잔기에 K48 연결 유비퀴틴화를 촉매하여 분해를 유도합니다. MARCHF2 결핍은 시험관 내 및 생체 내에서 선천성 면역 반응이 증가함에 따라 세균/바이러스 감염에 대한 현저한 저항성을 유발했습니다. 또한 MARCHF2(-/-) 마우스는 다량의 사이토카인 생성으로 인해 LPS 투여에 더 민감했습니다. 이러한 결과는 NEMO의 분자 조절에 대한 새로운 통찰력을 제공하며 선천성 면역 반응의 항상성 조절에서 MARCHF2의 중요한 역할을 시사합니다.[6]
Marchf2 녹아웃 마우스 생성
바이러스 또는 세균 감염 시 MARCHF2 단백질의 특정 면역 조절 기능을 탐구하기 위해 연구자들은 March2 녹아웃 마우스를 사용하여 MARCH2의 생리학적 역할을 규명했습니다. 연구 결과 MARCH2는 바이러스 또는 세균 감염 시 NF-κB 및 I형 IFN 신호 전달 경로의 음성 조절 인자임을 확인했습니다.
본 연구에서는 과배란 유도를 위해 4~6주령의 암컷 C57BL/6J 마우스를 배아 공여체로 사용했습니다. 교배 후 수정된 배아를 난관에서 채취하여 KSOM 배지에서 배양했습니다. 나리시기 미세주입 시스템이 장착된 올림푸스 IX71 도립 현미경을 사용하여 미세주입을 수행했습니다. gRNA(50 ng/μl)와 타겟 유전자 편집 mRNA(100 ng/μl)를 혼합하여 M2 배지 내에서 전핵이 관찰되는 접합체의 세포질에 주입했습니다. 주입된 배아는 37℃, 5% CO₂ 조건의 KSOM 배지에서 밤새 배양했습니다. 2세포기 단계의 배아를 가임신 ICR 암컷 마우스의 난관에 이식했습니다. 생후 3주령의 새끼 마우스에서 꼬리 샘플을 채취하여 게놈 DNA를 추출하고 시퀀싱했습니다. 예상되는 이형접합체 March2+/-를 교배했습니다. March2 KO 마우스를 스크리닝하기 위해 꼬리 생검에서 추출한 게놈 DNA 샘플을 사용하여 T7E1 분석을 수행했습니다. 간략히 설명하면 RNA 가이드 엔도뉴클레아제 표적 부위를 포함하는 게놈 영역을 네스티드 PCR 방법으로 증폭한 후 변성 및 재결합시켜 이중가닥 이형 DNA를 형성한 다음 T7 엔도뉴클레아제 1(New England Biolabs)로 처리하고 아가로스 겔 전기영동으로 분석했습니다. 이형 마우스를 제외한 후 동량의 야생형 PCR 산물을 혼합하여 추가 T7E1 분석을 수행하고 아가로스 겔 전기영동으로 WT와 KO 마우스를 분리하여(그림 1.G) March2 유전자 녹아웃을 확인했습니다.
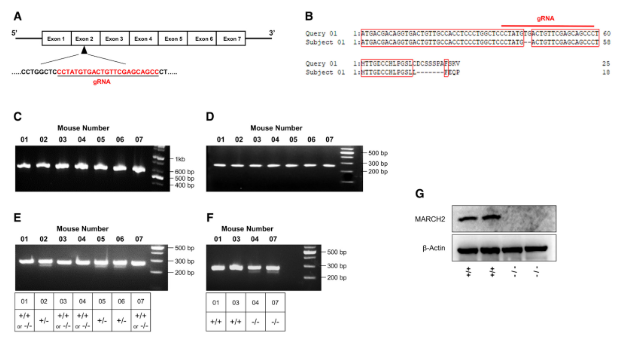
그림 1. MARCH2 녹아웃 마우스 생성 [6]
MARCHF2는 생체 내 숙주 방어에 중요한 역할을 합니다
선천성 면역 반응에서 MARCHF2의 잠재적 역할을 평가하기 위해 연구자들은 타겟 유전자 편집 시스템을 사용하여 C57BL/6 배경의 MARCHF2-/- 마우스를 생성하고 MARCH2 유전자의 유전적 녹아웃을 검증했습니다(그림 1A~G). MARCHF2-/- 마우스는 외관상 어떠한 이상도 나타내지 않았습니다.
먼저 생체 내 바이러스 감염에서 MARCHF2의 중요한 역할을 조사하기 위해 MARCHF2+/+ 및 MARCHF2-/- 마우스에 VSV-GFP를 정맥 주사했습니다(그림 2A). MARCHF2-/- 마우스의 혈청은 MARCHF2+/+ 마우스보다 복제 중인 바이러스가 더 적게 포함되어 있었습니다. 또한 연구자들이 혈청 내 사이토카인 양을 측정한 결과 MARCHF2-/- 마우스에서 IFN-β, IL-6, IL-12, TNF-α의 농도가 MARCHF2+/+ 마우스보다 더 높은 것을 확인했습니다(그림 2B). 나아가 바이러스 RNA 모방 리간드인 poly(I:C)를 주사한 마우스의 혈청 사이토카인 수준을 측정했습니다(그림 2C). 바이러스 감염에서 관찰된 바와 같이 리간드 자극 후 MARCHF2-/- 마우스의 혈청 사이토카인 수준이 MARCHF2+/+ 마우스보다 더 높았습니다. 이러한 결과는 MARCHF2가 생체 내 항바이러스 선천성 면역 반응에 관여함을 강력하게 시사합니다.
두 번째로 세균 감염에 대한 반응에서 MARCHF2가 역할을 하는지 조사하기 위해 연구자들은 MARCHF2+/+ 및 MARCHF2-/- 마우스에 치사량의 리스테리아 모노사이토제네스(Lm)를 투여했습니다. 그림 2D와 E에 나타난 바와 같이 MARCHF2-/- 마우스는 MARCHF2+/+ 마우스보다 세균 감염에 더 저항성이 있었습니다. 병원체 제거 효과를 확인하기 위해 집락 형성 단위(CFU) 적정 분석을 사용하여 비장과 간의 세균 부하를 조사했습니다(그림 2F와 G). 이 결과는 생존 분석 결과와 상관관계가 있었으며, MARCH2-/- 마우스의 세균 부하는 MARCHF2+/+ 마우스보다 유의하게 낮았습니다. 다음으로 MARCHF2-/- 마우스의 낮은 세균 부하가 이 동물의 높은 염증 반응 때문인지 조사했습니다. MARCH2-/- 마우스의 혈청, 비장, 간 샘플에는 MARCHF2+/+ 마우스보다 IL-6, IL-12, CCL5, CXCL10 수준이 더 높게 포함되어 있었습니다(그림 2H~J). 이러한 결과는 또한 MARCHF2가 생체 내 항세균 선천성 면역 반응에 관여하며, MARCH2 결핍이 항미생물 면역을 강화함을 나타냅니다.
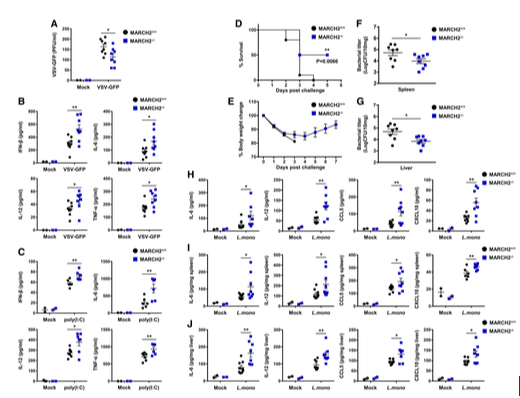
그림 2. MARCH2 결핍은 생체 내 항미생물 면역을 강화합니다 A, B. MARCH2+/+(n = 8) 또는 MARCH2-/-(n = 8) 마우스에 VSV-GFP(2 × 108 PFU/마우스)를 정맥 감염시키고 감염 후 12시간(hpi)에 혈청 샘플을 채취했습니다. (A) 플라크 분석으로 바이러스 역가를 분석했습니다. (B) 특정 ELISA로 IFN-β, IL-6, IL-12, TNF-α의 분비를 측정했습니다. C. MARCH2+/+(n = 6) 또는 MARCH2-/-(n = 6) 마우스에 poly(I:C)(200 μg/마우스)를 정맥 주사했습니다. 감염 후 2시간에 혈청 샘플을 채취하고 특정 ELISA로 IFN-β, IL-6, IL-12, TNF-α의 분비를 측정했습니다. D~J. MARCH2+/+(n = 8) 또는 MARCH2-/-(n = 8) 마우스에 1 × 106 CFU의 리스테리아 모노사이토제네스를 복강 내 투여했습니다. (D) 각 그룹의 마우스 생존율(D, 로그 순위 검정, **P < 0.01)과 체중 변화(E)를 나타냅니다. (F, G) 감염 후 3일(dpc)에 비장(F)과 간(G)의 세균 부하를 조사했습니다. (H~J) 감염 후 24시간에 각 그룹 마우스의 혈청(H), 비장(I), 간(J)에서 IL-6, IL-12, CCL5, CXCL10의 양을 측정했습니다. 데이터 정보: *P < 0.05, **P < 0.01 (양측 스튜던트 t-검정). 데이터는 평균 ± SEM으로 표시됩니다. [6]
세 번째로 MARCHF2가 리포다당류(LPS) 유도 패혈성 쇼크에 대한 감수성과 관련이 있는지 생체 내에서 조사하기 위해 MARCH2+/+ 및 MARCHF2-/- 마우스에 LPS를 복강 내 주사했습니다. 모든 MARCH2+/+ 마우스는 생존한 반면 MARCHF2-/- 마우스의 60% 이상이 사망했습니다(그림 3A와 B). 이러한 결과는 MARCHF2-/- 마우스가 MARCHF2+/+ 마우스보다 내독소 쇼크에 더 민감함을 시사합니다. 또한 비장 절편의 헤마톡실린-에오신 염색 결과 MARCHF2-/- 마우스에서 MARCHF2+/+ 마우스보다 염증 세포 침윤이 더 많이 관찰되었습니다(그림 3C). 따라서 연구자들은 MARCHF2가 LPS 매개 패혈성 쇼크 동안 염증 반응에 관여한다고 가설을 세웠습니다. 이를 검증하기 위해 ELISA를 사용하여 혈청 내 사이토카인 수준을 측정했습니다(그림 3D). 예상대로 MARCHF2-/- 마우스는 MARCHF2+/+ 마우스보다 염증 매개 인자의 발현이 현저히 높았습니다. 연구자들은 또한 비장과 간에서 전염증성 사이토카인/케모카인 유전자를 암호화하는 mRNA의 발현을 측정했습니다. 그림 3E와 F에 나타난 바와 같이 IL-6, TNF-α, CXCL10, IL-1β를 암호화하는 mRNA의 발현이 MARCHF2-/- 마우스에서 MARCHF2+/+ 마우스보다 유의하게 높았습니다. 이러한 데이터는 MARCHF2가 과도한 염증 조건에서 염증 반응을 약화시킴을 나타냅니다. 종합적으로 이러한 결과는 MARCHF2가 바이러스 및 세균 감염에 대한 선천성 면역 반응의 핵심 조절 인자임을 생체 내에서 증명합니다.
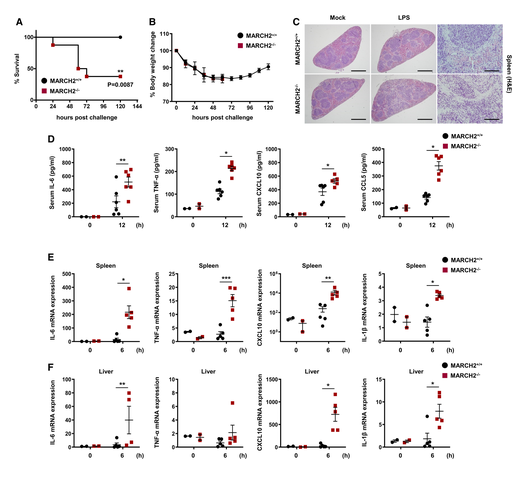
그림 3. MARCH2 결핍은 LPS 유도 패혈성 쇼크에 대한 감수성을 유도합니다 A, B. MARCH2+/+(n = 8) 또는 MARCH2-/-(n = 8) 마우스에 LPS(24 mg/kg)를 복강 내 투여했습니다. (A, B) 각 그룹의 마우스 생존율(A, 로그 순위 검정, **P < 0.01)과 체중 변화(B)를 나타냅니다. C. MARCH2+/+(n = 4) 또는 MARCH2-/-(n = 4) 마우스에 LPS(24 mg/kg)를 복강 내 투여했습니다. 각 그룹의 비장 절편 H&E 염색 대표 슬라이드를 나타냅니다. 스케일 바, 6 mm. D. MARCH2+/+(n = 6) 또는 MARCH2-/-(n = 6) 마우스에 LPS(24 mg/kg)를 복강 내 투여했습니다. 감염 후 12시간에 각 그룹 마우스의 혈청에서 IL-6, TNF-α, CXCL-10, CCL-5 수준을 ELISA로 측정했습니다. E, F. MARCH2+/+(n = 5) 또는 MARCH2-/-(n = 5) 마우스에 LPS(24 mg/kg)를 복강 내 투여했습니다. 마우스의 비장과 간에서 추출한 총 RNA로부터 cDNA를 제조했습니다. 감염 후 6시간에 각 그룹 마우스의 비장(E)과 간(F)에서 IL-6, TNF-α, CXCL10, IL-1β를 암호화하는 mRNA의 발현을 qPCR로 확인했습니다. 데이터 정보: *P < 0.05, **P < 0.01, ***P < 0.001 (양측 스튜던트 t-검정). 데이터는 평균 ± SEM으로 표시됩니다. [6]
요약: 향후 연구
설명된 바와 같이 Marchf2는 감염 후기 단계에서 NF-κB 조절 인자(NEMO)와 상호작용하고 염증 경로 관련 인자의 발현을 추가로 활성화합니다. Marchf2의 유전적 녹아웃은 세균 및 바이러스 감염에 대한 마우스의 저항성을 증가시키고 선천성 면역 반응을 강화했습니다. Marchf2가 바이러스 감염에서 중요한 역할을 하기 때문에, COVID-19 또는 향후 바이러스 유행에 대한 신약 개발에서도 잠재적 표적이 될 수 있을까요?
원스톱 마우스 모델 검색 플랫폼: MouseAtlas
MouseAtlas는 KO부터 인간화 마우스까지 유전자 및 제품 모델명 검색만으로 원하는 모델을 쉽게 찾을 수 있는 플랫폼입니다. 생체 마우스인지 정자 상태인지, 실시간 재고 상황, 검증 데이터, 상세 설명 등을 직관적으로 확인하고 바로 주문할 수 있습니다. 당사 내부 제품 관리 시스템과 연동되어 실시간으로 업데이트되며, 현재 39,000종 이상의 모델 마우스가 등록되어 있어 연구자들에게 매우 편리한 원스톱 솔루션을 제공합니다.
참고문헌
1.https://www.ncbi.nlm.nih.gov/gene/51257
2.https://www.uniprot.org/uniprot/Q9P0N8#function
3.https://alphafold.ebi.ac.uk/entry/Q9P0N8
4.https://www.genecards.org/cgi-bin/carddisp.pl?gene=MARCHF2&keywords=Marchf2
5.http://www.informatics.jax.org/marker/MGI:1925915
6.Chathuranga K, Kim TH, Lee H, Park JS, Kim JH, Chathuranga WAG, Ekanayaka P, Choi YJ, Lee CH, Kim CJ, Jung JU, Lee JS. Negative regulation of NEMO signaling by the ubiquitin E3 ligase Marchf2. EMBO J. 2020 Nov 2;39(21):e105139. doi: 10.15252/embj.2020105139. Epub 2020 Sep 16. PMID: 32935379; PMCID: PMC7604578.



